|
|

CELLS OF THE IMMUNE SYSTEM
T Cells and T Lymphopoiesis
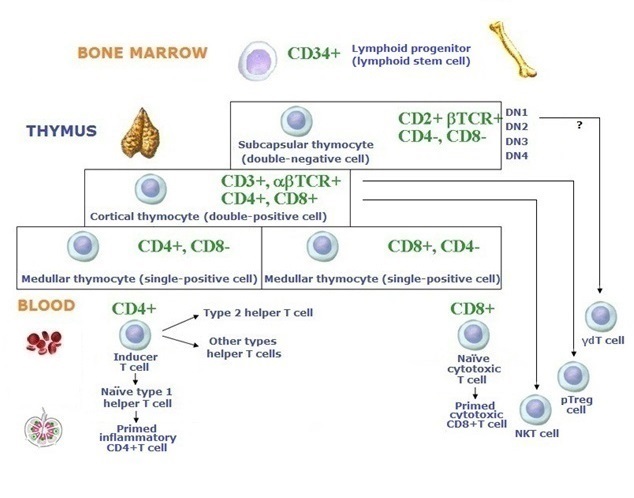
T cells are an important population of lymphocytes. From the 12th week of fetal life, thymocyte precursors originate from the lymphoid stem cell and migrate in response to chemokines via the blood from the bone marrow to the thymus. These cells have not yet rearranged their T-cell Receptor (TCR) genes, and they lack expression of TCR, accessory antigen receptor molecules, or coreceptors. At the 1st stage, in the subcapsular, or outer cortical zone, the thymocytes are double-negative (DN) cells, CD4- CD8-, but the β chain of TCR is expressed. For research purposes, these DN thymocytes are subdivided into DN1, DN2, DN3 and DN4 cells. At the 2nd stage, in the inner cortical zone thymocytes are double-positive (DP), CD4+CD8+, and have completed αβTCR. At the 3rd stage, they are single-positive (SP), CD4+ T cells or CD8+T cells.
The thymic microenvironment required for T lymphopoiesis is provided by giant epithelial "nurse" cells, macrophages, thymic dendritic cells and thymic NK cells. The T lymphopoiesis also depends on many cytokines and chemokines including IL-7.
At the DN stage, TCRβ genes begin rearranging first. At the DP stage, TCR α chain rearrangement occurs, and both chains are linked together. Gene products of Recombination Activating Genes (RAG-1 and RAG-2), expressed at all stages of T lymphopoiesis, are necessary for both β chain and α chain rearrangement.
It is believed that four major features of T lymphopoiesis or T-cell commitment are available.
into different clones by unique TCR (with a single antigenic specificity), which is linked to an accessory antigen receptor molecule, CD3, and one of two coreceptors, CD4+ or CD8+.
(1) The clonal division
(2) Two types of thymic selection:
positive selection, when TCRs have a low affinity for self-HLA molecules and self-antigens, and
negative selection (or clonal deletion), which induces apoptosis in thymocytes, which bind self-HLA molecules and self-antigens too well or do not bind them at all. However, a small portion of T cells can escape from apoptosis and come into an unresponsive state (clonal anergy). Positive selection leads to HLA restriction, and negative selection results in self-tolerance. About 95% of thymocytes cannot escape from negative selection. The gene AIRE upregulates negative selection.
(3) The division of all T cells into two subsets, naïve helper/inflammatory CD4+T cells, and naïve cytotoxic CD8+T cells. These subpopulations have different fates.
(4) The ability to form lifelong memory T cells after naïve T cells encounter antigens.
The remaining 5% of naïve T cells enter the bloodstream and lymph flow to differentiate into helper, effector or memory T cells. They have minimal cytoplasm, condensed chromatin, and little transcriptional activity. A phenotypic marker of naïve T cells is CD45RA+. The most recent coming from the thymus T cells are called recent thymic emigrants (RTE). Naïve T cells settle T dependent zones in the lymph nodes, periarteriolar sheaths in the spleen and parafollicular zones of MALT. Priming will start out when a T cell encounters an appropriate antigen. Naïve CD4+ and CD8+ T cells involved in adaptive immune responses become activated T cells, then effector T cells and memory T cells. Effector T cells appear to be the final stage of T-cell differentiation.
The natural killer (NKT) T cells similar to both T cells and NK cells, defined by their selective recognition of the CD1d HLA-like molecule and specialized to defense against cancerous cells make up about 1% of T cells in the peripheral blood. Peripheral regulatory FoxP3+ T (pTreg) cells function in suppressing immune responses by secretion of protolerogenic cytokines and apoptosis in effector cells. γδT cells, in part of the thymic development, called intraepithelial lymphocytes (IELs), protect the body against opportunistic microbes in the barrier organs.
In total, T cells account for 40-80% of peripheral lymphocytes in the blood.©V.V.Klimov