|
|

КЛЕТКИ ИММУННОЙ СИСТЕМЫ
Т-клетки и Т-лимфопоэз
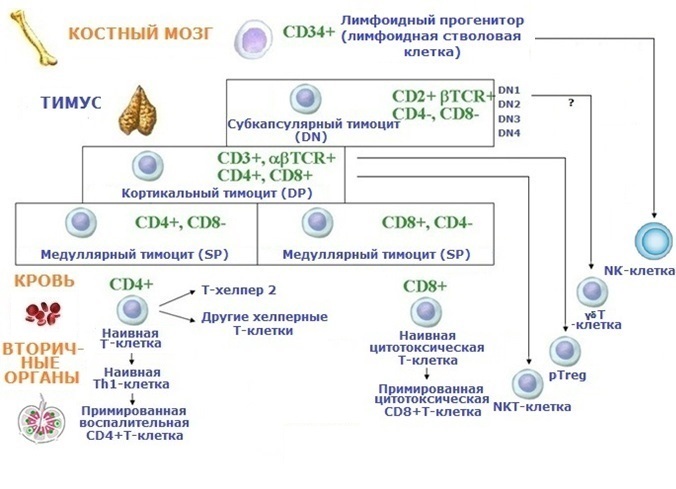
Антигеннезависимая дифференцировка (коммитмент) Т-клеток (T cells) начинается с их миграции в тимус (хоминг) в начале 12-й недели эмбриогенеза. В дальнейшем Т-лимфопоэз происходит, пока существуют даже самые маленькие эпителиально-макрофагальные островки в тимусе, но на ранних стадиях онтогенеза он протекает очень интенсивно. Тимоциты проходят последовательно стадию "двойных негативных клеток" (double-negative cells, DN), "двойных позитивных клеток (double-positive cells, DP) и "монопозитивных клеток" (single-positive cells, SP). Первоначально на стадии DN происходит реаранжировка TCRβ-генов, затем на стадии DP - TCRα-генов, и обе цепи соединяются вместе. Генные продукты - ферменты RAG-1 и RAG-2 (Recombination Activating Genes) экспрессирутся на всех стадиях Т-лимфопоэза, т.к. они необходимы для генетических реаранжировок в тимоцитах. Важнейшим цитокином (ростовым фактором) для Т-лимфопоэза является IL-7.
Основные черты Т-лимфопоэза:
1. Экспрессия специфического TCR (как маркёра принадлежности к определённому Т-клону), ассоциированных молекул CD3 и корецепторов CD4 или CD8.
2. Селекция («экзамен») в ходе дифференцировки:
- положительная селекция - отбор и выпуск в кровоток клонов, способных к распознаванию собственных HLA и экзоантигенов;
- отрицательная селекция - делеция путём апоптоза клонов с высокоаффинными TCRs, направленными против собственных антигенов (до 95% тимоцитов).
3. Разделение на субпопуляции CD4+ и CD8+.
4. Приобретение Т-клетками способности к долгосрочной иммунной памяти после адаптивных иммунных ответов.
Полнота отрицательной селекции определяется геном AIRE (autoimmune regulator), от чего зависит риск развития аутоиммунных расстройств. Некоторые Т-лимфоциты в ходе коммитмента приобретают не один, а два отличающихся по специфичности TCR.
По окончанию Т-лимфопоэза до 5% тимоцитов избегают апоптоза, составляя около 50 миллионов T-клеточных клонов. Наивные Т-клетки мигрируют в Т-зависимые зоны периферических органов (паракортикальные зоны лимфатических узлов, периартериальные пространства селезёнки, околофолликулярные области МАЛТ) для потенциального участия в aнтигензависимой дифференцировке (прайминге). Вновь пришедшие из тимуса Т-клетки получили название "недавние тимические эмигранты (recent thymic emigrants, RTE)". Миграция (хоминг) Т-клеток контролируется хемокинами CXCL13 (BLC), CCL19 (ELC) и CCL21 (SLC). Каждый Т-клон "ждёт" случая, когда антиген, к которому он программирован (коммитирован), попадёт в организм. Некоторые Т-клоны, возможно, до конца жизни остаются без «работы». T-клетки c αβTCR составляют 40-80 % среди лимфоцитов периферической крови.
γδТ-клетки (с γδTCR-типом рецептора) являются минорной популяцией в крови (0,1-0,5%); они частично тимического происхождения и присутствуют в значительном количестве в барьерных органах. Большинство этих клеток имеет особенный корецептор - CD8αα (в отличие от CD8αβ у αβTСR-лимфоцитов). Для них не характерны разнообразие специфичностей и клональная экспансия, и они сразу вырабатывают специфические эффекторные молекулы. Мишенями для них являются оппортунистические патогены.
Ещё одна субпопуляция - NKT-клетки (natural killer T cells) - по фенотипу похожи одновременно на Т-клетки и NK-клетки. Они способны распознавать гликолипидные антигены, загруженные на молекулы CD1d, подобные молекулам HLA. Они необходимы в противоопухолевом иммунитете и процессах толерантности.
Около 0,05% T-клеток становятся nTreg-клетками (non-adaptive natural regulatory T cells), ответственными за аутотолерантность в тимусе. Однако самыми важными клетками для поддержания иммунной толерантности на периферии являются периферические антиген-специфические FoxP3+рTreg-клетки (adaptive peripheral regulatory T cells), дифференцирующиеся из наивных Т-клеток. Возникают и другие регуляторные Т-клетки: FoxP3-Tr1 и FoxP3-Th3.©В.В.Климов